Viruses, first observed in honeybees, have been documented in bumblebees and the prevalence and load of some RNA viruses have been associated with managed honeybees. Shared flowers may be the bridge across which viruses pass between bees but no study has yet demonstrated that bumblebees can pick up viruses while foraging on contaminated flowers. Here, through a series of mechanistic laboratory experiments and mathematical modeling, we ask whether viruses can be transmitted between bee genera on shared flowers and how transmission can be effectively mitigated. We demonstrated that deformed wing virus (DWV) can be transmitted from infected honeybees to bumblebees through the use of shared red clover. We were also able to show that the route may work in reverse and bumblebees could contribute to the spread as well. Our model showed that reducing vector-mediated transmission in honeybee colonies could potentially lead to a far greater reduction in bumblebee infection than simply reducing the number of honeybees. Additionally, we identified a dilution effect, whereby increasing floral abundance reduced transmission. Synthesis and applications. In this study, we showed that DWV may be spread between bee genera through the shared use of flowers. Through mathematical simulation, we identified two practical management options for reducing spread. The combination of treating honeybees effectively for the Varroa mite, a known vector of DWV, and increasing floral abundance where honeybees and native pollinators share the landscape were shown to reduce the spread of DWV within bee communities in simulations.

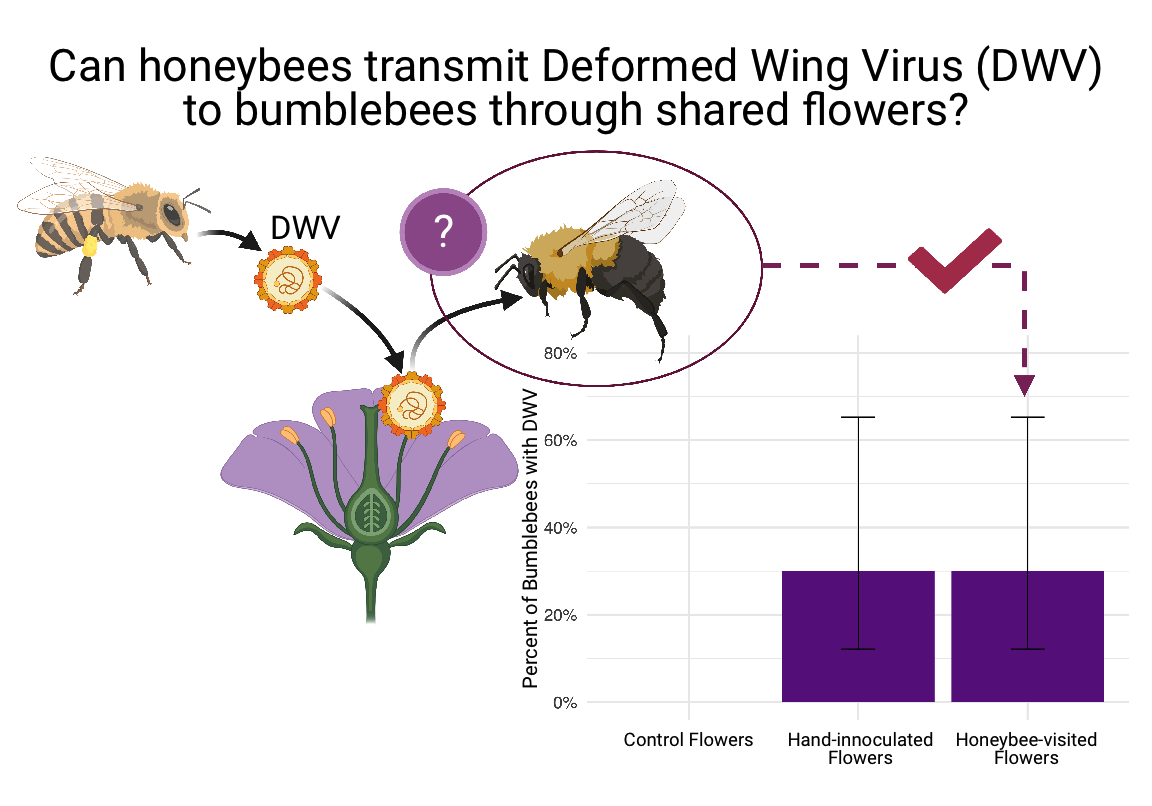
RNA viruses, once considered specific to honey bees, are suspected of spilling over from managed bees into wild pollinators; however, transmission routes are largely unknown. A widely accepted yet untested hypothesis states that flowers serve as bridges in the transmission of viruses between bees. Here, using a series of controlled experiments with captive bee colonies, we examined the role of flowers in bee virus transmission. We first examined if honey bees deposit viruses on flowers and whether bumble bees become infected after visiting contaminated flowers. We then examined whether plant species differ in their propensity to harbor viruses and if bee visitation rates increase the likelihood of virus deposition on flowers. Our experiment demonstrated, for the first time, that honey bees deposit viruses on flowers. However, the two viruses we examined, black queen cell virus (BQCV) and deformed wing virus (DWV), were not equally distributed across plant species, suggesting that differences in floral traits, virus ecology and/or foraging behavior may mediate the likelihood of deposition. Bumble bees did not become infected after visiting flowers previously visited by honey bees suggesting that transmission via flowers may be a rare occurrence and contingent on multiplicative factors and probabilities such as infectivity of virus strain across bee species, immunocompetence, virus virulence, virus load, and the probability a bumble bee will contact a virus particle on a flower. Our study is among the first to experimentally examine the role of flowers in bee virus transmission and uncovers promising avenues for future research.

The decline of many bumblebee species (Bombus spp.) has been linked to an increased prevalence of pathogens caused by spillover from managed bees. Although poorly understood, RNA viruses are suspected of moving from managed honeybees (Apis mellifera) into wild bumblebees through shared floral resources. We examined if RNA viruses spillover from managed honeybees, the extent to which viruses are replicating within bumblebees, and the role of flowers in transmission. Prevalence and active infections of deformed wing virus (DWV) were higher in bumblebees collected near apiaries and when neighboring honeybees had high infection levels. We found no DWV in bumblebees where honeybee foragers and honeybee apiaries were absent. The prevalence of black queen cell virus (BQCV) was also higher in bumblebees collected near apiaries. Furthermore, we detected viruses on 19% of flowers, all of which were collected within apiaries. Our results corroborate the hypothesis that viruses are spilling over from managed honeybees to wild bumblebees and that flowers may be an important route for transmission.

Honey bees are important pollinators of agricultural crops and the dramatic losses of honey bee colonies have risen to a level of international concern. Potential contributors to such losses include pesticide exposure, lack of floral resources and parasites and pathogens. The damaging effects of all of these may be exacerbated by apicultural practices. To meet the pollination demand of US crops, bees are transported to areas of high pollination demand throughout the year. Compared to stationary colonies, risk of parasitism and infectious disease may be greater for migratory bees than those that remain in a single location, although this has not been experimentally established. Here, we conducted a manipulative experiment to test whether viral pathogen and parasite loads increase as a result of colonies being transported for pollination of a major US crop, California almonds. We also tested if they subsequently transmit those diseases to stationary colonies upon return to their home apiaries. Colonies started with equivalent numbers of bees, however migratory colonies returned with fewer bees compared to stationary colonies and this difference remained one month later. Migratory colonies returned with higher black queen cell virus loads than stationary colonies, but loads were similar between groups one month later. Colonies exposed to migratory bees experienced a greater increase of deformed wing virus prevalence and load compared to the isolated group. The three groups had similar infestations of Varroa mites upon return of the migratory colonies. However, one month later, mite loads in migratory colonies were significantly lower compared to the other groups, possibly because of lower number of host bees. Our study demonstrates that migratory pollination practices has varying health effects for honey bee colonies. Further research is necessary to clarify how migratory pollination practices influence the disease dynamics of honey bee diseases we describe here.